 Syno®C 引物合成
Syno®C 引物合成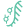 RNA合成
RNA合成 mRNA合成
mRNA合成 Syno®GS 基因合成
Syno®GS 基因合成 载体构建
载体构建 高通量及DNA文库构建
高通量及DNA文库构建 CRISPR基因编辑平台
CRISPR基因编辑平台 病毒包装
病毒包装 基因测序与组学分析
基因测序与组学分析 重组蛋白表达平台
重组蛋白表达平台 抗体工程平台
抗体工程平台 多肽服务
多肽服务 生物信息学分析与设计
生物信息学分析与设计 CRISPR文库
CRISPR文库 合成生物学产品
合成生物学产品 ProXpress蛋白快速检测
ProXpress蛋白快速检测 CRISPR 质粒
CRISPR 质粒
CRISPRi跨菌株“失效”?从实验室株到临床大肠杆菌的遗传设计规则重构
2026年,ACS Synthetic Biology 上发表了一项系统性研究,对基于 dCas9 与 dCas12a 的 CRISPRi 技术在不同大肠杆菌菌株间的可迁移性进行了全面评估。研究者提出了“宿主特异性设计规则”——包括系统选择策略、毒性对照设置以及双 gRNA 协同优化方案,为非模式菌株的基因调控工具箱构建提供了可量化的设计准则。

研究背景与科学问题的提出
合成生物学与功能基因组学研究长期依赖于模式菌株 Escherichia coli K-12(如 MG1655)建立的遗传工具箱。CRISPR干扰(CRISPRi)技术凭借其可编程性与转录抑制的高效性,已成为细菌基因调控的核心工具。然而,当研究者试图将成熟的CRISPRi系统从实验室模式菌株迁移至非模式菌株(如临床分离株、益生菌株)时,往往面临宿主特异性限制(Host-specific limitations)的严峻挑战。
本研究聚焦于一个核心的科学问题:基于II型(dCas9)与V型(dCas12a)的CRISPRi系统在系统发育相近但生态位迥异的大肠杆菌菌株中,其转录抑制效率与细胞毒性是否存在显著的宿主依赖性差异?
实验设计与方法学
研究团队构建了一个高度标准化的评估框架,旨在消除质粒拷贝数、启动子强度等变量干扰,实现跨菌株的精准比对。
1. 菌株与系统选择
宿主菌株:
涵盖实验室株(MG1655, Phylogroup A)、益生菌株(Nissle 1917, EcN, Phylogroup B2)、尿路致病株(CFT073, Phylogroup B2; UMN026, Phylogroup D)。
CRISPRi系统:
①Sp-dCas9(S. pyogenes):经典的II型系统,PAM = NGG。
② Fn-dCas12a(F. novicida):V型系统,PAM = TTTV,具有pre-crRNA处理能力。
③Lb-dCas12a(Lachnospiraceae ND2006):另一种V型系统,PAM = TTTV。
2. 遗传回路设计
采用双质粒、双诱导系统:
• 质粒1(调控质粒):携带dCas效应蛋白与gRNA,分别受 PTet(aTc诱导)和 PTac(IPTG诱导)控制,呈背向方式排列并辅以强终止子,以最大限度降低泄漏表达。
• 质粒2(报告质粒):携带荧光报告基因(eYFP用于dCas9,sfGFP用于dCas12a)。
3. 定量表征
• 抑制效率:通过流式细胞术(Flow Cytometry)进行单细胞水平检测,计算相对表达单位(REU)及抑制倍数(Fold Repression)。
• 细胞毒性:通过微孔板读数仪监测不同诱导浓度下的生长曲线,计算特定生长速率(Specific Growth Rate)与OD600变化。
核心结果与深度解析
gRNA设计规则的保守性与菌株特异性漂移
研究发现,尽管不同菌株间的gRNA抑制倍数存在相关性(Pearson’s r = 0.889–0.975),但绝对抑制指呈现显著的菌株依赖性。
Sp-dCas9:在CFT073中表现最佳(17.6-fold),而在EcN中显著下降(7-fold)。值得注意的是,UMN026表现出极高的本底荧光(Uninduced REU: 5.3–11.1),暗示该菌株可能存在更强的转录泄漏或报告基因稳定性差异。
dCas12a系统:Fn-dCas12a在所有测试菌株中均表现出优于Sp-dCas9的抑制深度(MG1655最高达93-fold),且gRNA活性相关性更强(r > 0.96)。
宿主特异性的细胞毒性图谱
这是本研究的重要发现之一:dCas蛋白的表达对宿主生长的影响并不遵循系统发育树。
Sp-dCas9:在UMN026中表现出极端的毒性(4 ng/mL aTc诱导下OD降低94%,生长速率降低72%),且存在显著的泄漏毒性(Uninduced condition)。
Lb-dCas12a:总体毒性最低,特别是在UMN026中表现出比Fn-dCas12a更好的相容性。
关键结论:dCas蛋白的毒性并非单纯由启动子输出量决定(Figure S8),提示不同菌株的蛋白折叠负荷、膜通透性差异或内源性CRISPR-Cas系统互作可能是导致毒性差异的潜在机制。
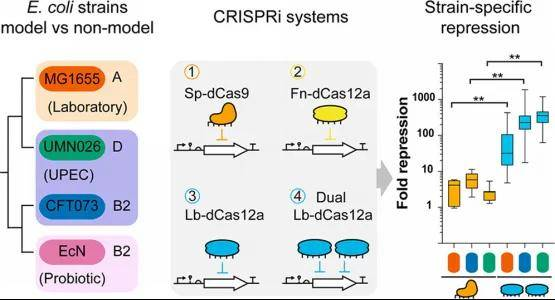
双gRNA阵列策略的构建与优化
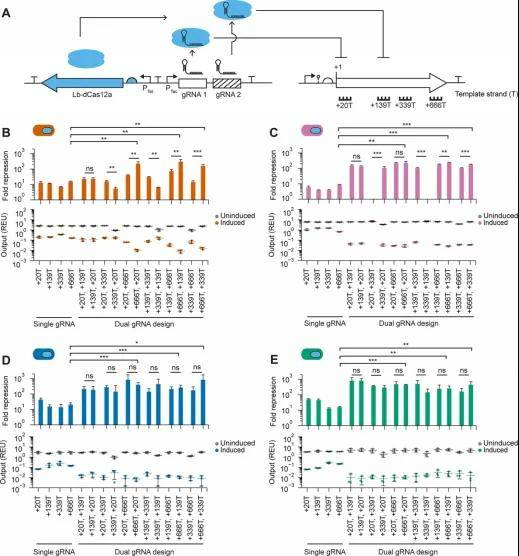
针对单gRNA在部分临床菌株中抑制不足的问题,研究团队开发了基于Lb-dCas12a的双gRNA串联阵列(Multiplexed Dual gRNA Array),靶向sfGFP的CDS区域。
机制洞察:
非线性协同效应:双gRNA的抑制效果既不符合简单的加性模型(Additive model),也不完全符合乘性模型(Multiplicative model),表明存在复杂的空间位阻协同或染色质结构重塑效应。
位置效应(Positional Effects):在MG1655和EcN中,gRNA在阵列中的顺序显著影响抑制效率(如将低效gRNA置于5’端会拖累下游加工),而在UPEC菌株中这种依赖性较弱。这反映了CRISPR阵列的加工动力学(Processing kinetics)具有宿主特异性。
讨论与设计准则
本研究为CRISPRi在非模式菌株中的应用提供了严谨的设计指南:
1.gRNA可迁移,但抑制倍数需重调:靶向非模板链启动子区或CDS区的gRNA设计规则在不同菌株间高度保守,但具体的抑制倍数必须重新标定。
2.系统选择策略:若追求低毒性与多重编辑潜力,Lb-dCas12a是临床菌株(尤其是UPEC)的首选;若需极致抑制,推荐采用双gRNA阵列策略。
3.警惕“隐性毒性”:在将CRISPRi系统迁移至新菌株时,必须设立严格的dCas-only对照组(无gRNA),以区分生长抑制是由靶基因下调引起,还是由dCas蛋白本身的寄生负担(Parasitic burden)导致。
方法学亮点
对于有志于开发非模式菌株遗传工具的研究者,本文补充材料中的以下技术细节尤为值得关注:
• 电转化条件优化:针对CFT073和UMN026等难以转化的临床株,详细描述了使用无盐LB培养基(No-salt LB)及低OD600收获窗口(0.3–0.4)的电感受态制备方案。
• gRNA设计脚本:作者开源了基于Python的gRNA设计流程,整合了Bad Seed效应过滤与脱靶分析(Cas-OFFinder),适用于不同CRISPRi系统的自动化设计。
• 动力学建模:文中尝试使用简化动力学模型预测低表达水平下的抑制动态,为理性调节dCas与gRNA的表达剂量提供了理论框架。
该研究打破了“同一物种内CRISPRi工具通用”的经验主义假设,通过系统生物学手段揭示了遗传元件与宿主背景互作的复杂性,为临床菌株的精准基因调控奠定了坚实基础。
针对此类非模式菌株遗传工具开发中的挑战,泓迅生物依托其独特的“GPS”平台,能够为研究者提供从序列智能设计到基因编辑的一体化解决方案。欢迎咨询交流!

扫码立即咨询
Ban, H., Rondthaler, S. N., Lebovich, M., et al. (2026). Cross-Strain Transferability of CRISPRi Systems and Design Rules from Laboratory to Clinical Escherichia coli Strains. ACS Synthetic Biology, 15(7), 1993–2010.
https://doi.org/10.1021/acssynbio.6c00075


























